
RESEARCH OVERVIEW
Kristen L. Kroll, Ph.D., Professor
Developmental, Regenerative, and Stem Cell Biology Program, Molecular Cell Biology Program
Our research focuses on defining gene regulatory networks (GRNs) that control neural cell specification, neurogenesis, and the generation of specific neuronal cell types. We are particularly interested in understanding how epigenetic regulation modulates these networks and how their dysregulation contributes to neurodevelopmental disorders and birth defects, including epilepsy, autism spectrum disorder, and neural tube defects. This work uses directed differentiation of human pluripotent stem cells (embryonic stem cells and induced pluripotent stem cells), mouse models, and a wide range of cellular, molecular, and genomic approaches, to define roles for transcriptional and epigenetic regulation in shaping developmental transitions.
The lab has three main research interests, outlined below:
For more information see Cellular Modeling of Neurodevelopmental Disorders.
A major focus of our work is to identify transcriptional and epigenetic regulatory controls underlying the development of human cortical interneurons and to understand how their dysregulation contributes to neurodevelopmental disorders. The mammalian cerebral cortex consists of two major types of neurons, excitatory glutamatergic projection neurons that convey information to different regions of the brain and GABAergic interneurons that provide local inhibitory inputs to modulate responses of projection neurons and prevent over-excitation. Imbalances between excitatory and inhibitory neuronal activities can emerge during cortical development. These imbalances often reduce inhibitory signaling through dysfunctional specification, migration, differentiation, or survival of interneurons. Interneuron hypoplasia or dysfunction contributes to many neurological disorders, including epilepsy, schizophrenia, autism spectrum disorder, and intellectual disability syndromes.
We are modeling gene regulatory networks that control human cortical interneuron specification and differentiation at a genome-wide level by generating mature interneurons from human pluripotent stem cells (embryonic stem cells (hESCs) and induced pluripotent stem cells (iPSCs)).

Human pluripotent stem cell-derived cortical interneurons (hcINs): electrophysiology and engraftment into the murine adult and neonatal brain. (A) Scheme: Synapsin (SYN)-GFP expressing hcINs (after PSA-NCAM selection; scale bars=100µM) were transplanted into the (B) hippocampus (CA3) of adult mice (B’=boxed region in B; scale bar=50µM) or (C) cortex of post-natal 2 NOD/SCID mice (C’=boxed region in C, scale bar=100µM). B-C: 30 days post-transplantation shown. The ‘scan slide’ mode in the Nikon confocal software was used to assemble a composite view of the whole brain slice from several individual images. (D) Scheme: SYN-GFP expressing hcINs (after PSA-NCAM selection; scale bar=100µM) were co-cultured on rat cortical astrocyte feeders for 3 wks. prior to electrophysiology. (E) Sub- and supra-threshold stimulation by injection of depolarizing current (52 and 56 pA for 40 msec). (F) Multiple action potentials during injection of depolarizing current (56 pA for 800 msec). (G) Spontaneous inhibitory postsynaptic currents (sIPSCS) recorded at -80 and -20 mV are inhibited by exposure to 100 µM bicuculline methiodide (black bars). (H) Whole-cell currents recorded during a voltage step from -80 to +50 mV in control (black trace) and 0.5 µM tetrodotoxin (TTX, red trace). (I) Currents evoked by exposure to 100 µM kainate (K), NMDA, GABA, and glycine (gly). (J) Evoked IPSC elicited by a presynaptic voltage step. (K) Electrophysiological properties of hESC-derived hcINs (mean +/- SEM).
We are integrating genome-wide binding profiles of key transcription factors and chromatin regulators, developmental transcriptome and epigenome changes, and effects of manipulating these activities. This work has revealed new regulators of interneuron development, including transcriptional and chromatin modifying activities and non-coding RNAs, enriched in and/or involved in interneuron specification and differentiation.

Integrating genome-wide binding profile of the transcription factor NKX2-1 and enriched gene expression in hPSC-derived interneuron progenitors (hMGEs) and neurons (hcINs). (A) RNA-seq data for cortical interneurons. k-means clustered data, based upon expression z-scores. (B-C) NKX2-1-associated genes with enriched expression at d15 (B) or d35 (C)
We have also defined genes with interneuron-enriched expression whose mutation contributes to human neurodevelopmental disorders, including inherited epilepsies and autism spectrum disorder. In collaboration with the WU Intellectual and Developmental Disability Research Center (IDDRC), we are deriving iPSCs from epilepsy and autism patients with mutations in some of these genes and are performing directed differentiation into cortical excitatory and inhibitory neurons and cerebral organoids. These are subjected to a battery of assays to define developmental, cellular, molecular, and functional abnormalities that contribute to the disorder. We can use these models to assess the relative contributions of genetic background versus pathogenic mutations, by comparisons of isogenic neurons with versus without engineered correction of mutations. We are also using these models to assess how differential expressivity of a disorder among family members carrying a known pathogenic mutation manifests in cellular, molecular, and functional differences between neurons from these family members.

Requirements for the epilepsy and IDD gene, CHD2, in cortical interneuron development. (A) Differentially regulated genes upon CHD2 knockdown (KD), with some highlighted. (B) Genes with reduced expression in the CHD2 KD RNA-seq were also diminished in hMGEs with biallelic CHD2 KO vs wild-type hMGEs. (H-I) CHD2 KO reduced TUBB3 and HCN4 immunopositive cells and (J) neurite length. (K) Currents mediated by TTX-sensitive sodium channels during steps from a holding potential of -80 mV to test potentials ranging from -60 to +35 mV. Traces were obtained by subtracting currents recorded during exposure to TTX from currents recorded in control external solution without TTX. Peak inward current is plotted as a function of test potential for wild type (black) and knock out (red) cells. (L) Phase plots of action potentials from 13 wild type (grey) and 9 knock out (red) neurons. The initial increase in dV/dt occurs at more negative membrane potentials in knock out cells, indicating a difference in action potential threshold. Inset at top right plotted on expanded scales, lower right the action potentials are aligned at the point of maximal upslope. For B, I-J, p-values defined by unpaired student’s t-test, *<0.05, **<0.01, ***<0.001. In D, H, scale bar=50µM.
Neural cell fate acquisition is mediated by transcription factors expressed in nascent neuroectoderm, including Geminin and members of the Zic transcription factor family. In our prior work, we identified chromatin association profiles for Geminin and Zic1 during neural fate acquisition at a genome-wide level (Sankar et al., 2016). We determined how Geminin deficiency affected histone acetylation at gene promoters during this process. We integrated these data to determine that Geminin associates with and promotes histone acetylation at neurodevelopmental genes, while Geminin and Zic1 bind a shared gene subset. Geminin- and Zic1-associated genes exhibit embryonic nervous system-enriched expression and encode other regulators of neural development. Both Geminin and Zic1-associated peaks are enriched for Zic1 consensus binding motifs, while Zic1-bound peaks are also enriched for Sox3 motifs, suggesting co-regulatory potential. Accordingly, we found that Geminin and Zic1 could cooperatively activate the expression of several shared targets encoding transcription factors that control neurogenesis, neural plate patterning, and neuronal differentiation. We used these data to construct gene regulatory networks underlying neural cell fate acquisition. Establishment of this molecular program in nascent neuroectoderm directly links early neural cell fate acquisition with regulatory control of later neurodevelopment.
We are characterizing how disruption of this network contributes to the etiology of neural tube defects (NTDs) and other neurodevelopmental disorders. Although NTDs are the second most common birth defect (~1:1000 fetuses and newborns), their causation is poorly understood. This is, in part, because NTD occurrence usually has a multi-factorial etiology, with both genetic and environmental susceptibility factors contributing to phenotypic manifestation and severity. Therefore, defining aspects of this regulatory network that are dysregulated to contribute to NTDs in multiple models may identify core processes that, when disrupted, increase NTD susceptibility. This work also defined novel epigenetic regulators with likely roles in neural development, some of which we are functionally assessing in ongoing work.

Figure Legend. Genome-wide binding profiles for Geminin and Zic1 were used to define gene regulatory networks in neural cell fate acquisition. (A-D) Comparison of Gmnn-associated and Zic1-associated genes in NE. (A) A subset of genes is associated with both Gmnn and Zic1. p-value (Chi-square test with Yates’ correction) < 2.2X10-16. (B) z-scores for enrichment of expression in ES versus CNS tissues for all Zic and/or Gmnn associated genes. (C) Comparison of all Gmnn or Zic1 associated genes, subsets that undergo Gmnn-dependent acetylation, and transcription factors, with their relative enrichment of expression in embryonic CNS tissues. (D) GO enrichment analysis for associated genes with increased versus decreased expression in embryonic CNS, relative to ES cells. (E) Gene regulatory networks in neural cell fate acquisition. Gmnn- and Zic1-associated genes in NE that encode transcription factors and epigenetic regulatory activities were assessed for co-association with Sox2 and Sox3 in ES-derived NE (see text). The set of these genes that exhibit CNS-enriched expression (>two-fold greater expression in E14 CNS, relative to ES cells), and were associated with Gmnn and/or Zic1 plus Sox2 and/or Sox3 were used to build gene regulatory networks showing associations.

Neurite extension assay: control versus Autism Spectrum Disorder (ASD) affected proband-derived neurons 
Analysis of synaptic punctae in control versus Autism Spectrum Disorder (ASD) affected proband-derived neurons 
Comparison of gene expression profiles of neurons from an unaffected mother and her son, the Autism Spectrum Disorder (ASD) affected proband 
Differentially regulated genes in the control versus Autism Spectrum Disorder (ASD) affected proband-derived neurons include a network involved in neuronal migration 
Interneurons (red) and excitatory neurons (green) were cocultured as neuroids to measure migration of interneurons. Comparison of migration in an unaffected mother (UM) and her son, the Autism Spectrum Disorder (ASD) affected proband 
A human embryonic stem cell colony 
Neuronal differentiation of hPSC-derived neural progenitors 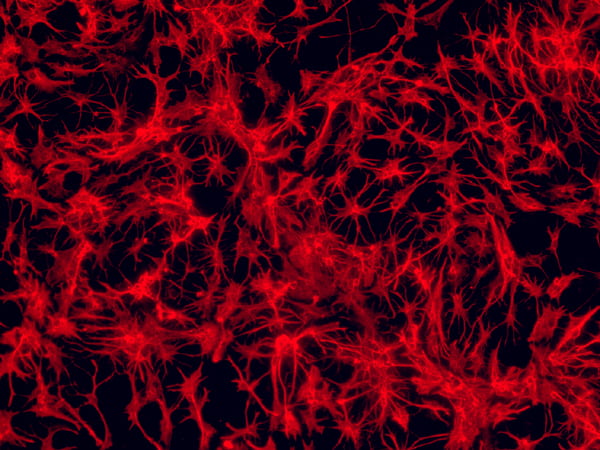
Differentiation of mouse embryonic stem cells into astrocytes (GFAP staining) 
Regionally restricted expression of molecular markers in the mouse embryonic neural tube 
Cortical interneurons differentiated from human pluripotent stem cells